Какие бактерии образуют споры. Споры и спорообразование
Существуют некоторые виды бактерий, которые производят тела круглой или овальной формы, отличающиеся сильным светопреломлением. Эти образования носят название эндоспор. Спорообразование является одной из стадий цикла развития определенных микроорганизмов в ответ на неблагоприятное воздействие внешней среды, выработанное в процессе эволюции в борьбе за сохранение вида. Недостаток питательных веществ вызывает у некоторых микроорганизмов различные реакции, обеспечивающие подготовку клетки к длительному периоду, в течение которого питательные вещества недоступны. Переход к спорообразованию наблюдается при истощении питательного субстрата, при недостатке углерода, азота или фосфора, изменении рН среды и т.д. Спорообразование присуще, в основном, палочковидным микроорганизмам (бациллы и клостридии, и сравнительно редко наблюдается у кокков (Sarcina urea, Sarcina lutea) и извитых форм (Desulfovibrio desulfuricans).
Спорообразование происходит во внешней среде, на питательных средах и не наблюдается в тканях человека и животных. Пpoцесс спорообразования разделяется на семь последовательных стадий, характеризующихся различными цитологическими изменениями (рис. 12).
Подготовительные стадии (стадии 0 и I). На этих стадиях в клетке еще нет морфологически видимых изменений, но уменьшается количество воды и уплотняется цитоплазма.
Стадия проспоры (стадия II) является первой стадией спо-руляции, распознаваемой морфологически. Она характеризуется появлением проспоровой перегородки, которая делит клетку на маленькую проспору и большую материнскую клетку. Это ключевой этап споруляции.
В течение стадии поглощения проспоры (стадия III) происходит пространственное обособление маленькой проспоры, которая переходит в цитоплазму материнской клетки. Снаружи проспоры образуется двойная мембранная структура.
Стадия предспоры характеризуется образованием кортекса (плотной споровой оболочки) внутри мембранной структуры проспоры (стадия IV) и конденсацией на его поверхности белков (стадия V).
На стадии созревания (стадия VI) споровая оболочка получает дальнейшее развитие и становится устойчивой к химическим агентам и нагреванию. Сформировавшаяся спора занимает примерно 1/10 часть материнской клетки.
Итоговой стадией является освобождение зрелой споры от материнской клетки (стадия VII). Процесс образования споры протекает в течение 18-20 часов.
Вследствие наличия плотной многослойной оболочки, имеющей пластинчатое строение, минимального количества воды и высокого содержания кальция, липидов и дипиколиновой кислоты, споры обладают большой устойчивостью к действию факторов окружающей среды и дезинфицирующих средств. Они выдерживают относительно высокие и низкие температуры, длительное высушивание, действие радиации, токсичных веществ и т.д. Они могут десятилетиями сохраняться в неблагоприятных условиях.
Попадая в благоприятные условия, споры прорастают и вновь превращаются в вегетативные формы. Процесс прорастания спор начинается с поглощения воды. Они набухают, увеличиваются в размерах. Из оболочки на полюсе, в центре или между полюсом и центром появляется отросток, из которого вытягивается палочка. Процесс прорастания спор идет значительно быстрее и занимает 4 - 5 часов.
По характеру локализации в теле микроорганизмов споры располагаются:
1. Центрально (палочка сибирской язвы, aнтракоидная палочка и др.).
2. Субтерминально - ближе к концу (возбудитель ботулизма и др.).
3. Терминально - на конце палочки (возбудитель столбняка).
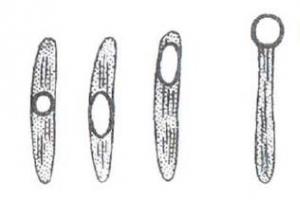
У отдельных видов спорообразующих микроорганизмов диаметр спор превышает поперечник бактериальной клетки. Если споры локализуются субтерминально, такие бактерии принимают форму веретена. К ним принадлежат клостридии маслянокислого брожения. У некоторых клостридии, например, у возбудителя столбняка, споры располагаются терминально, их клетка напоминает барабанную палочку (рис. 13).

Рис. 13. Формы и расположение спор в бациллах.
Способность к спорообразованию используют в систематике микробов, а также при выборе методов обеззараживания предметов, помещений, пищевых продуктов, различных изделий.
СПОРЫ (греч. spora сеяние, посев) - микроскопические зачатки, образуемые нек-рыми растениями и бактериями при бесполом размножении.
Споры растений
С. образуют растения различных систематических групп - водоросли, грибы, многие виды высших растении.
Библиография: Вилли К. А. и Детье В. Д ж. Биология (Биологические процессы и законы), пер. с англ., с. 267, 660, М., 1974; Жизнь растений, под ред. А. А. Федорова, т. 1 - 4, М., 1974 - 1978; Корн М. Я. и Солов ь-е в H. Н. Об использовании фазовоконтрастной микроскопии для наблюдения спор в окрашенных мазках, Лаборат. дело, № 6, с. 51, 1961; Краткий определитель бактерий Берги, под ред. Дж. Хоулта, пер. с англ., с. 286, М., 1980; Работ-н о в а И. Л. Общая микробиология, с. 65, М., 1966; Руководство по микробиологической диагностике инфекционных болезней, под ред. К. И. Матвеева, с. 27, М., 1973; Стейниер Р., Э д е л ь б e р г Э. и Ингрэм Д ж. Мир микробов, пер. с англ., т. 3, с. 184, М., 1979; The bacterial spore, ed. by G. W. Gould a. A. Hurst, L.-N. Y., 1969.
Т. В. Вещикова; М. Я. Корн (бакт.); М. Я. Корн и М. Ф. Култаев (цветн. рис.).
Спорообразование начинается когда клетка испытывает недостаток питат вещ-в или в среде накопилось большое кол-во продуктов обмена. Тогда клетка выделяет до 40% воды, цитоплазма уплотняется. В центре образуется дипикколиновая кислота. У молодых клеток этой кислоты нет. Эта кислота взаимод с ионами кальция и образует комплекс дипиколинат кальция, который обеспечивает споре состояние покоя и термоустойчивость. Образ-ся проспора. пространство между обол материнской мембраны и проспорой заполнено пептидогликаном, из которого формируется стенка споры. После созревания споры клетка разруш-ся и спора выходит наружу, она переносит все неблагопр.усл. спора имеет тройную обол. Попадая в благопр. усл, спора поглощает воду, набухает, дипиколинат кальция выходит наружу. Наруж обол лопается, во внеш среду выходит трубка – проросток. Она дает начало новой клетке.из 1 споры выходит 1 клетка. Существуют клетки устойчивые к неблагопр усл, но спор не образуют. Это покоящиеся клетки – цисты. Споры у бактерий служат только для сохранения и выживания организма в неблагоприятных условиях (тогда как у грибов, например, споры служат еще и для размножения).
Культивирование.
Размножение микроорг-в осущ-ся на питат средах. Посев культуры микробов на такие среды проводят микробной петлей и пастеровскими пипетками.
Различают два основных способа культивирования микроорганизмов – периодическое и непрерывное. При периодическом культивировании клетки помещают в закрытый сосуд определенного объема, содержащий питательную среду, и задают начальные условия. Постепенно увеличивается плотность популяции, снижается концентрация питательных веществ и накапливаются продукты обмена, т.е. условия существования микроорганизмов изменяются. Непрерывное (проточное) культивирование позволяет зафиксировать культуру в какой-то определенной фазе (обычно экспоненциальной). При этом состав среды и условия роста остаются постоянными. Этого добиваются постоянным прибавлением новой питательной среды в сосуд для выращивания и одновременным
удалением такого же количества среды с клетками.
Посев проводят в пробирках на мпб и мпа, а также смесь культур на мпа в чашках Петри.
1 в чашки Петри вносят питат среду, для этого мпа расплавляют на водяной бане. Содержимое пробирок выливают над пламенем горелки и вращат движениями распределяют на поверхности дна чашки. Чашку переворачивают дном вверх и ставят в термостат.
СПОСОБЫ ОПРЕДЕЛЕНИЯ АНАЭРОБНЫХ БАКТЕРИЙ.
Анаэробы растут без свободного кислорода, т.к. образующийся пероксид водорода окисляет цитоплазму и вызывает гибель микробной клетки. Создание условий с пониженным сод-ем кислорода в возд достиг-ся разными методами.
Физич метод - анаэробиоз создается путем мех-го удаления воздуха, что достигается вакуум-масляным насосом. Химический метод основан на поглощении кислорода хим-ми вещ-ми. Био метод – аэробов и анаэробов выращивают вместе. Для этого плотную питат среду мпа в чашке петри делят на 2 части. На одну из них высевают аэробов, на др – анаэробов. После заливки краев чашки парафином ее ставят в термостат. Комбинированный метод. Анаэробиоз создается путем сочетания неск методов. На принципе этого метода основано исп-ие среды Кита-Тароцци – на дно пробирки помещают кусочки отварной печени, а сверху заливают маслом.
Могут возникать: из целой клетки, накопившей питательные вещества и утолщившей оболочку (экзоспоры многих сине-зеленых водорослей). при делении Протопласта на большое число спор (эндоспоры некоторых сине-зеленых водорослей, рис. 1, 1). в результате уплотнения и сжатия протопласта внутри оболочки клетки и образования поверх него новой многослойной оболочки (у бактерий). при распаде особых участков мицелия на членики (у актиномицетов, рис. 1, 2). У растений — эукариотов, обладающих типичными ядрами, имеющих 3 основных типа спор (оо-, мито- и мейоспоры) и занимающих разное место в циклах развития, могут быть соответственно и 3 варианта С.: ооспорогенез, митоспорогенез и мейоспорогенез. Обычно под С. понимают образование мейоспор (мейоспорогенез). Ооспорогенез связан с процессом оплодотворения и, следовательно, со сменой ядерных фаз в циклах развития. заканчивается образованием ооспор (у многих зелёных водорослей и оомицетов), ауксоспор (у диатомей), зигоспор (у зигомицетов), представляющих собой одноядерные или многоядерные зиготы.
Митоспорогенез приводит к возникновению митоспор, формирующихся по нескольку или в обльшом числе в результате митотических делений (см. Митоз) гаплоидных [например, зооспоры ряда водорослей (рис. 1, 3) и грибов], реже диплоидных (например, карпоспоры большинства флоридей) клеток или без делений — моноспоры эдогониума (рис. 1, 4), бангиевых, немалионовых. к смене ядерных фаз не приводит. Протекает в одноклеточных митоспорангиях (например, в зооспорангиях улотрикса, моноспорангиях эдогониума, цистокарпиях флоридей), а одноклеточные водоросли как бы сами становятся спорангиями (рис. 1, 5).
Митоспорогенез может наблюдаться при распадении мицелия, состоящего из клеток, содержащих дикарионы, например у головнёвых и ржавчинных грибов . Мейоспорогенез связан со сменой диплофазы в циклах развития как низших, так и высших растений гаплофазой. У низших растений мейоспоры возникают в результате Мейоза или вскоре после него из митотически разделившихся гаплоидных клеток, образовавшихся при мейозе. У водорослей и грибов с гаплоидным циклом развития С. происходит при прорастании зиготы (ооспоры), диплоидное ядро которой, делясь мейотически, образует 4 гаплоидных ядра. при этом возникают 4 мейоспоры (например, зооспоры хламидомонад, рис. 1, 6, апланоспоры улотрикса), либо 3 из четырёх гаплоидных ядер отмирают и образуется лишь 1 мейоспора (например, у спирогиры, рис. 1, 7), либо за мейозом следует 1-3 митотических деления и формируются 8-32 споры (например, у бангиевых).
У водорослей, имеющих изоморфный и гетероморфный циклы развития, мейоспорогенез протекает в одноклеточных мейоспорангиях и характеризуется образованием либо 4 мейоспор (например, тетраспоры бурых водорослей и большинства флоридей, рис. 1, 8), либо 16-128 мейоспор (например, зооспоры ламинариевых, рис. 1, 9) вследствие 2-5 митотических делений, следующих за мейозом. В спорангиях сумчатых грибов (сумках, или асках) возникшие в результате мейоза 4 гаплоидных ядра делятся митотически и формируются 8 эндогенных мейоспор (аскоспор). В базидиях (спороносных органах) базидиальных грибов после мейоза возникают по 4 гаплоидных ядра, которые перемещаются в специальные выросты на поверхности базидий. в дальнейшем эти выросты с гаплоидными ядрами, т. и. базидиоспоры, отделяются от базидий (рис. 1, 10). Высшие растения образуют только мейоспоры, мейоспорогенез протекает в многоклеточных спорангиях. Обычно в результате митотических делений диплоидных клеток археспория возникают т. н. спороциты (мейотически делящиеся клетки), формирующие по 4 споры (тетрады спор).
Равноспоровые папоротникообразные продуцируют морфологически и физиологически одинаковые споры (рис. 2, 1), из которых развиваются обоеполые заростки. У разноспоровых папоротникообразных и семенных растений осуществляются микро- и мегаспорогенез, мейоспорогенез, т. е. возникают споры двух типов. Микроспорогенез происходит в микроспорангиях и завершается образованием большого числа микроспор (рис. 2, 2), прорастающих затем в мужские заростки. мегаспорогенез — в мегаспорангиях, где в меньшем числе — часто даже 4 или 1 — созревают мегаспоры (рис. 2, 3), прорастающие в женские заростки.
Развивающиеся спороциты и споры (у большинства высших растений) питаются веществами, получаемыми из клеток тапетума (слоя, выстилающего изнутри полость спорангия). У многих растений клетки этого слоя, расплываясь, образуют периплазмодий (протоплазматическую массу с дегенерирующими ядрами), в котором оказываются спороциты, а затем и споры. У некоторых растений в формировании периплазмодия участвует и часть спороцитов. В мегаспорангиях (семезачатках) некоторых покрытосеменных в результате мейоза образуются клетки с 2 или 4 гаплоидными ядрами, соответствующие 2 (рис. 2, 4) или 4 (рис. 2, 5) мегаспорам. из этих клеток развиваются женские гаметофиты — т. н. биспорические и тетраспорические зародышевые мешки. О С. у простейших см. в ст. Споры.
Лит.: Мейер К. И., Размножение растений, М., 1937. Курсанов Л.И., Комарницкий Н. А., Курс низших растений, М., 1945. Магешвари П., Эмбриология покрытосеменных, пер. с англ., М., 1954. Тахтаджян А. Л., Высшие растения, т. 1, М. — Л., 1956. Поддубная-Арнольди В,А., Общая эмбриология покрытосеменных растений, М., 1964: Smith G. М., Cryptogamic botany, 2 ed., v. 1-2, N. Y. — L., 1955. Lehrbuch der Botanik f
ьr Hochschulen, 29 Aufl., Jena, 1967.
А. Н. Сладков.
Рис. 1. Спорообразование у низших растений. 1 — образование и выход эндоспор у сине-зеленой водоросли Dermocarpa. 2 — распадение мицелия на членики у актиномицета Nocardia. 3 —
Споры – своебразная форма покоящихся фирмикутных бактерий, т. е. бактерий с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium – веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Процесс образования спор проходит ряд последовательных стадий:
подготовительная. Изменяется метаболизм, завершаетется репликация ДНК и происходит ее конденсация. Клетка содержит два или более нуклеоида, один из них локализуется в спорогенной зоне, остальные - в цитоплазме спорангия. Одновременно синтезируется дипиколиновая кислота;
стадия предспоры. Со стороны цитоплазматической мембраны вегетативной клетки происходит врастание двойной мембраны, или септы, отделяющей нуклеоид с участком уплотненной цитоплазмы (спорогенная зона). В результате чего образуется проспора, окруженная двумя мембранами;
образование оболочек. Вначале между мембранами про-споры образуется зачаточный пептидогликановый слой, затем над ним откладывается толстый пептидогликановый слой кор-текса и вокруг его наружной мембраны формируется споровая оболочка;
созревание споры. Заканчивается образование всех структур споры, она становится термоустойчивой, приобретает характерную форму и занимает определенное положение в клетке.
15. Сложные методы окраски. Окраска по Граму, её этапы.
Последовательно нанести на препарат определенные красители, различающиеся по химическом составу и цвету, протравы, спирты, кислоту и др. Это позволяет выявиться стркутуры клеток и дифференцировать одни виды микроорганизмов от других.
Окраска по Граму.
1. На фиксированный мазок на препарат нанести карболово-спиртовый раствор генцианового фиолетового через полоску фильтровальной бумаги. Через 1-2 мин ее снять, а краситель смыть.
2. Нанести раствор люголя на 1-2 мин
3. Обесцветить этиловым спиртом в течении 30-6- с до прекращения отхождения фиолетовых струек красителя.
4. Промыть водой.
5. Докрасить водным раствором фуксина 1-2 мин, промыть, высушить и микроскопировать. Грам +: темно-фиолетовый цвет. Грам -:красные
16. Окраска по Цилю-Нильсену и ее этапы.
Метод окраски по Цилю - Нельсену - метод окраски микроорганизмов для выявления кислотоустойчивых микобактерий (возбудителей туберкулёза, микобактериозов, лепры), актиномицетов и других кислотоустойчивых микроорганизмов. Кислотоустойчивость микроорганизмов обусловлена наличием в их клетках липидов, воска и оксикислот. Такие микроорганизмы плохо окрашиваются разведёнными растворами красителей. Для облегчения проникновения красителя в клетки микроорганизмов нанесённый на препарат карболовый фуксин Циля подогревают над пламенем горелки. Окрашенные микроорганизмы не обесцвечиваются слабыми растворами минеральных кислот и спирта.
Фиксированный на пламени горелки мазок окрашивают в течение 3 – 5 мин. раствором карболового фуксина Циля или окрашенной фуксином бумажкой с подогреванием до появления паров, но не доводя краситель до кипения.
Дают препарату остыть, бумажку снимают, сливают избыток красителя, препарат промывают водой.
Окрашенный препарат обесцвечивают 5%-ным раствором серной кислоты в течение 3 – 5 с или 96° этиловым спиртом, содержащим 3% по объему хлористоводородной кислоты, несколько раз погружая стекло с мазком в стаканчик с солянокислым спиртом.
После обесцвечивания остаток кислоты сливают и тщательно промывают препарат водой.
Докрашивают дополнительно метиленовым синим Леффлера 3 – 5 мин.
Окрашенный препарат промывают водой, подсушивают и микроскопируют.
При окраске препаратов кислоустойчивые бактерии окрашиваются фуксином в рубиново-красный цвет и не обесцвечиваются кислотой.
Некислотоустойчивые бактерии, а также элементы ткани и лейкоциты под действием кислоты обесцвечиваются и приобретают цвет дополнительного красителя.